
Monday, December 1, 2014
Tree of Chordates
Using a combination of anatomical, molecular, and fossil evidence, biologists have developed hypotheses for the evolution for chordate groups. The first transition was the development of a head that consists of a brain at the anterior end of the dorsal nerve cord, eyes and other sensory organs, and a skull. These innovations opened up a completely new way of feeding for chordates: active predation. All chordates with a head are called craniates. The origin of a backbone came next. The vertebrates are distinguished by a more extensive skull and a backbone, or a vertebral column, composed of a serious of bones called vertebrae. These skeletal elements enclose the main parts of the nervous system. The skull forms a case for the brain, and the vertebrae enclose the nerve cord. The vertebrae skeleton is an endoskeleton, made of either flexible cartilage or a combination of hard bone and cartilage. Bone and cartilage are mostly nonliving material. But because there are living cells that secrete the nonliving material, the endoskeleton can grow with the animal. The next major transition was the origin of jaws, which opened up new feeding opportunities. The evolution of lungs or lung derivatives, followed by muscular lobed fins with skeletal support, opened the possibility of life on land. Tetrapods, jawed vertebrates with two pairs of limbs, were the first vertebrates on land. The evolution of amniotes, tetrapods with a terrestrially adapted egg, completed the transition to land. -Biology book page 390
{kind=link}
From Ovule to Seed in a Gynosperm
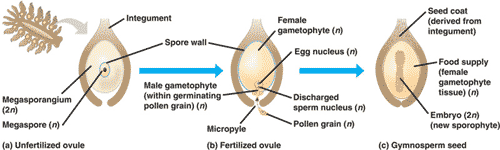
The evolution of vascular tissue solved the terrestrial problems of supporting the plant body and obtaining water and minerals from the soil. However, the challenges of reproduction and dispersing offspring on dry land remained. In contrast to seedless plants, which produce flagellated sperm that need moisture to reach an egg, seed plants--gymnosperms and angiosperms--have pollen grains that carry their sperm producing cells through the air. In addition, the offspring of seedless plants are sent off into the world as haploid, single celled spores that must survive independently as gametophytes before producing the next sporophyte generation. Seed plants launch next generation sporophytes that are ready to grow. In seed plants, a specialized structure within the sporophyte houses all reproductive stages, including spores, eggs, sperm, zygotes, and embryos. In gymnosperms such as pines and other conifers, this structure is called a cone. Cones are modified shoots that serve a reproductive function. -Biology book page 348-349
{kind=link}
Ulva
Green algae, which is named for their grass-green chloroplasts, include unicellular and colonial species as well as multicellular seaweeds. Ulva, or sea lettuce, is a multicellular green algae. Like many multicellular algae and all land plants, Ulva has a complex life cycle that includes an alternation of generations. In this type of life cycle, a multicellular diploid form alternates with a multicellular haploid form. Multicellular diploid forms are called sporophytes, because they produce spores. The sporophyte generation alternates with a haploid generation that features a multicellular haploid form called a gametophyte, which produces gametes. In Ulva, the gametophyte and sporophyte organisms are identical in appearance, although they differ in chromosome number. The haploid gametophyte produces gametes by mitosis, and fusion of the gametes begins the sporophyte generation. In turn, cells in the sporophyte undergo meiosis and produce haploid, flagellated spores. The life cycle is completed when a spore settles to the bottom of the ocean and develops into a gametophyte. -Biology book page 336

Bacteria and Archaea
Researchers recently discovered that many prokaryotes once classified as bacteria are actually more closely related to eukaryotes and belong in a domain of their own. As a result, prokaryotes are now classified in two domains: Bacteria and Archaea. Many bacterial and archaeal genomes have now been sequenced. When compared with each other and with the genomes of eukaryotes, these genome sequences strongly support the three-domain view of life. Some genes of archaea are similar to bacterial genes, others are eukaryotic genes, and still others seem to be unique to archaea. Differences between the rRNA sequences provided the first clues of a deep division among prokaryotes. Other differences in the cellular machinery for gene expression include differences in RNA polymerases and in the presence of introns within genes. The cell walls and membranes of bacteria and archaea are also distinctive. Bacterial cell walls contain peptidoglycan, while archaea do not. Furthermore, the lipids forming the backbone of plasma membranes between the two domains. Intriguingly, archaea have at least as much in common with eukaryotes as they do with bacteria. -Biology book page 325


{kind=link}
{kind=link}
Three Prokaryotes
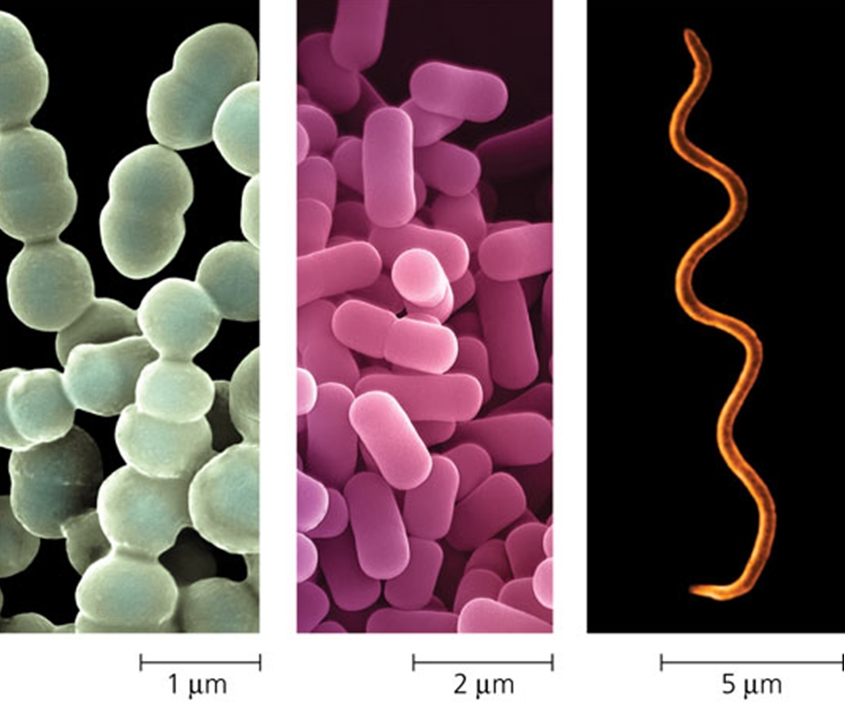
Some of the diversity of prokaryotes is evident in their external features, including shape, cell walls, and projections such as flagella. These features are useful for identifying prokaryotes as well as helping the organisms survive in their environments, Determining cell shape by microscopic examination is an important step in identifying prokaryotes. Spherical prokaryotic cells are called cocci. Rod-shaped prokaryotes are called bacilli. A third prokaryotic cell shape is spiral, like a corkscrew. Spiral prokaryotes that are relatively short and rigid are called spirilla; those with longer, more flexible cells that cause Lyme disease, are called spirochetes. Nearly all prokaryotes have a cell wall, a feature that enables them to live in a wide range of environments. The cell wall provides physical protection and prevents the cell from bursting in a hypotonic environment. Some prokaryotes have external structures that extend beyond the cell wall. Many bacteria and archaea are equipped with flagella, adaptions that enable them to move about in response to chemical or physical signals in their environment. Biology book page 320-321

{kind=link}
Protistan Diversity
Protists are bewilderingly different. Oxygen-using prokaryotes established residence within other, larger cells. These endosymbionts evolved into mitochondria, giving rise to heterotrophic eukaryotes. Autotrophic eukaryotes also arose through endosymbiosis of a prokaryote by a eukaryote after a heterotrophic eukaryote engulfed an autotrophic cynobacterium. If the cynobacterium continued to function within its host cell, its photosynthesis would have provided a steady source of food for the heterotrophic host and thus given it a significant selective advantage. And because the cynobacterium had its own DNA, it could reproduce to make multiple copies of itself within the host cell. In addition, cynobacterium could be passed on when the host reproduced. Over time, the descendants of the origin cynobacterium evolved into chloroplasts. The chloroplast-bearing lineage of eukaryotes later diversified into the autotrophs green algae and red algae. On subsequent occasions during eukaryotic evolution green algae and red algae themselves became endosymbionts following ingestion by different heterotrophic eukaryotes. The heterotrophic host cells enclosed the algal cells in food vacuoles but the algae survived and became cellular organelles. The presence of the endosymbionts, which also had the ability to replicate themselves, gave their hosts a selective advantage. -Biology book page 331

{kind=link}
Subscribe to:
Comments (Atom)